Universal Chalcidoidea Database
Notes on families
Eurytomidae
Main diagnostic characters
1. Pronotum, viewed from above, with sides more or less parallel and forming a subrectangular collar (95%)
2. Antenna with fewer than 13 segments (100%)
3. Body robust to elongate, about 1.4-6.0mm in length (100%)
4. Body strongly sculptured and black (80%)
5. Mesoscutum with notauli deep and complete (100%)
Included taxa
The family currently includes 88 genera and 1424 species placed in 3 subfamilies as follows: Eurytominae (73/1366), Heimbrinae (2/7), Rileyinae (12/50), unplaced (1/1).
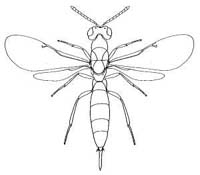 |
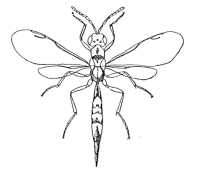 |
| Eurytominae (Axanthosoma sp.) |
Eurytominae (Aximogastra bahiae) |
Biology
The Eurytomidae contains species which exhibit a wide range of biologies, but the majority seem to be endophytic, either as phytophages or as parasitoids of phytophagous insects. There are two large groups of phytophagous eurytomids in Britain, those that develop on endosperm in seeds, and those that feed in plant stems, especially stems of grasses. The seed-feeding group is represented by two taxa, Systole, species of which feed in the seeds of Umbelliferae (Claridge, 1959b), and Bruchophagus, many species of which develop in the seeds of Leguminosae. The stem-mining group is represented in Britain by the large genus Tetramesa. Claridge (1961b) reviewed the biology of a number of British species. Several develop in the central cavity of grass stems, feeding above the nodes. They may be solitary or gregarious, but there is no external sign of their presence, and their effect on the flowering head is usually slight. Other species produce obvious stem-galls, and these often result in stunting of the flower head. In the United States several species of Tetramesa are pests of cereal crops, and a number of British seed-feeding eurytomids (eg Bruchophagus gibbus which attacks white clover, Trifolium repens) can assume pest status abroad (Valentine, 1970).
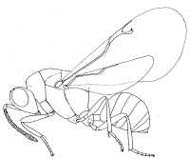 |
 |
| Eurytominae (Bruchophagous acaciae) |
Eurytominae (Eurytoma sp.) |
The majority of species of Eurytoma and Sycophila are entomophagous for at least part of their larval development, though several are known to complete their feeding on plant tissue (Varley, 1937). Most of these entomophagous species are idiobiont ectoparasitoids of insect larvae feeding within plant tissue. Hosts attacked include Coleoptera, gall-forming Hymenoptera (mostly Cynipinae), Diptera (especially Tephritidae) and Lepidoptera (Claridge, 1959a; Claridge & Askew, 1960). Fisher (1970) described the biology and life history of Eurytoma curculionum, a common larval parasitoid of an Apion weevil mining stems of docks (Rumex spp.). This eurytomid may develop either as a primary parasitoid of Apion, or as a facultative hyperparasitoid on other ectoparasitoids (eg Chlorocytus sp.) attacking the weevil larva. The eurytomid female paralyses a mature host larva before depositing an egg on or near it. The larva of another British species, Eurytoma rosae, is predaceous in multi-chambered galls on rose made by the cynipine genus Diplolepis. The eurytomid chews its way from cell to cell consuming several cynipine larvae in succession (Blair, 1944). Considerable polyphagy is shown by another eurytomid which inhabits cynipid galls. This species, Eurytoma brunniventris, may parasitize a cynipid gall maker, its Synergus inquiline, other chalcid parasitoids, or even feed on the gall tissue (Askew, 1961a). Not all entomophagous species of Eurytoma are associated with phytophagous insects. For example, E. rubicola is a common parasitoid of sphecids nesting in bramble-stems, ovipositing through the stem and developing as an idiobiont ectoparasitoid of the prepupa (Danks, 1971).
 |
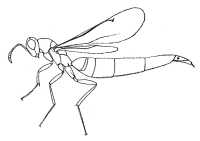 |
| Eurytominae (Tetramesa sp.) |
Rileyinae (Macrorileya oecanthi) |
A few eurytomids that attack cecidogenic insects are endoparasitic. Sycophila biguttata is found in Britain associated with oak galls. It develops as an endoparasitoid of the cecidogenic cynipid larva (Askew, 1984). Eurytoma serratulae develops as a koinobiont endoparasitoid of a tephritid that causes galls on thistles (Cirsium spp.). The eurytomid oviposits into an early instar larva, which continues to develop normally until late summer. Unparasitized tephritids overwinter as mature larvae, but parasitized tephritids attempt to pupate. The contents of the puparium are consumed by the parasitoid larva, which then overwinters in situ (Claridge, 1961a).
Most eurytomids are solitary parasitoids and, even though more than one egg may be laid on a host, larval competition ensures that only a single parasitoid develops. However, a few Eurytoma species are known which are gregarious ectoparasitoids of Lepidoptera (eg Piel, 1933). One of these, an Oriental species, is of particular interest as it is an obligate multiparasitoid (cleptoparasitoid). It can only oviposit on its host, a mature lepidopterous larva in a thick cocoon, if a chrysidid first bites an oviposition hole in the cocoon. The eurytomid oviposits through the plugged chrysidid hole.
A few eurytomids are recorded as predators of eggs of other insects (Clausen, 1940). Many of these belong to the non-British subfamily, the Rileyinae.
The eurytomid egg is very characteristic with a short process at the micropylar end and a long flattened filamentous process at the opposite end. Many that are deposited externally are clothed with short spines, and the form of these spines has been useful for separating sibling species (Claridge & Askew, 1960; Fisher, 1965). The first instar larva is hymenopteriform, oval to elongate, and each of its segments may have sensory setae. There are usually at least five pairs of spiracles present. The mature larva of endoparasitic species is typically hymenopteriform, but those of phytophagous species are more slender (see Askew, 1971: fig. 65). The comparative morphology of the final instar larvae of five species of eurytomid was studied by Roskam (1982). Most species overwinter as mature larvae.
Identification
Claridge 1959a, b; 1961c (British species of some genera); Peck, Boucek & Hoffer, 1964 (Central European genera); Ferriere, 1950 (most European species of Eurytoma and Bruchophagus); Narendran, 1994 (Indian genera and species), Szelényi, 1976a, b (Palaearctic species of Eurytoma and Bruchophagus); Nikol'skaya, 1952 (Russian species of Eurytoma and Bruchophagus); Boucek, 1988 (Australasian genera).
References
Askew, R.R. 1961. Eupelmus urozonus Dalman (Hym. Chalcidoidea) as a parasite in cynipid oak galls. Entomologist 94:196-201.
Askew, R.R. 1971. Parasitic insects :xvii+316pp. American Elsevier Publishing Co., Inc., New York .
Askew, R.R. 1984. The biology of gall wasps :223-271. In: Ananthakrishnan, T.N. (Editor) Biology of gall insects Oxford & IBH Publishing Co., New Delhi, Bombay and Calcutta
Blair, K.G. 1944. A note on the economy of the rose bedeguar gall, Rhodites rosae L. Proc. S. Lond. ent. nat. Hist. Soc. 1943-44:55-59.
Boucek, Z. 1988. Australasian Chalcidoidea (Hymenoptera). A biosystematic revision of genera of fourteen families, with a reclassification of species. :832pp.. CAB International, Wallingford, Oxon, U.K., Cambrian News Ltd; Aberystwyth, Wales.
Claridge, M.F. 1959a. A contribution of the biology and taxonomy of the British Species of the genus Eudecatoma Ashmead (Hym., Eurytomidae). Transactions for the Society of British Entomology 13:149-168.
Claridge, M.F. 1959b. Notes on the genus Systole Walker, including a previously undescribed species (Hym., Eurytomidae). Entomologist's Monthly Magazine 95:38-43.
Claridge, M.F. 1961a. Biological observations on some eurytomid (Hymenoptera: Chalcidodea) parasites associated with Compositae, and some taxonomic implications. Proceedings of the Royal Entomological Society of London (A) 36:153-158.
Claridge, M.F. 1961b. A contribution to the biology and taxonomy of some Palaearctic species of Tetramesa Walker (=Isosoma Walk.; =Harmolita Motsch.) (Hymenoptera: Eurytomidae) with particular reference .to the British fauna. Transactions of the Entomological Society of London 113:175-216.
Claridge, M.F. 1961c. An advance towards a natural classification of eurytomid genera (Hym., Chalcidoidea) with particular reference to British forms. Transactions of the Society for British Entomology 14:167-185.
Claridge, M.F.; Askew, R.R. 1960. Sibling species in the Eurytoma rosae group (Hym., Eurytomidae). Entomophaga 5:141-153.
Clausen, C.P. 1940. Entomophagous Insects :688pp. McGraw Hill, New York; London.
Danks, H.V. 1971. Biology of some stem-nesting aculeate Hymenoptera. Transactions of the Royal Entomological Society of London 122:323-395.
Ferriére, C. 1950. Notes sur les Eurytoma (Hym., Chalcidoidea). I. les types de Thomson et de Mayr. Mitt. Schweiz. Ent. Ges. 23:377-410.
Fisher, J.P. 1965. A contribution to the biology of Eurytoma curculionum Mayr. (Hym., Eurytomidae.). Entomophaga 10:317-318.
Narendran, T.C. 1994. Torymidae and Eurytomidae of Indian subcontinent. 500pp. University of Calicut, Kerala, India.
Nikol'skaya, M. 1952. Chalcids of the fauna of the USSR (Chalcidoidea). Opredeliteli po Faune SSSR, Izdavaemie Zoologicheskim Institutom Akademii Nauk SSR 44:575pp. Akademiya Nauk SSSR, Moscow and Leningrad. (In Russian; English translation: 1963: Israeli Prog. for Scient. Transl., Jerusalem 593pp)
Peck, O.; Boucek, Z.; Hoffer, A. 1964. Keys to the Chalcidoidea of Czechoslovakia (Insecta: Hymenoptera). Memoirs of the Entomological Society of Canada No 34:170pp, 289 figs.
Piel, O. 1933. Monema flavescens Wkr. and its parasites (Lepidoptera, Heterogeneidae). Lingnan Sci. J. 12(supplement):173-202.
Roskam, J.C. 1982. Larval characters of some eurytomid species (Hymenoptera, Chalcidoidea). Proc. Kon. ned. akad. Wetensch. 85:293-305.
Szelényi, G. 1976a. Mongolian eurytomids (Hymenoptera: Chalcidoidea). II. Acta Zool. Acad. Sci. Hung. 22:173-187.
Szelényi, G. 1976b. Mongolian eurytomids (Hymenoptera: Chalcidoidea). III. Acta Zool. Acad. Sci. Hung. 22:397-405.
Valentine, E.W. 1970. A list of the phytophagous Hymenoptera in New Zealand. New Zealand Entomologist 4:52-62.
Varley, G.C. 1937. Description of the eggs and larvae of four species of chalcidoid Hymenoptera parasitic on the knapweed gall-fly. Proceedings of the Royal Entomological Society of London (B) 6:122-130, 5 figs.
Previous page | Next pageLast updated 07-Jun-2004 Dr B R Pitkin